| Table of Contents | |
|
Review Article
| ||||||
| Embryology, comparative anatomy, and congenital malformations of the gastrointestinal tract | ||||||
| Melinda Danowitz1, Nikos Solounias2 | ||||||
|
1B.A, Medical Student, Anatomy, New York Institute of Technology College of Osteopathic Medicine, Old Westbury, NY, USA.
2PhD, Professor, Anatomy, New York Institute of Technology College of Osteopathic Medicine, Old Westbury, NY, USA. | ||||||
| ||||||
|
[HTML Abstract]
[PDF Full Text]
[Print This Article]
[Similar article in Pumed] [Similar article in Google Scholar] |
| How to cite this article |
| Danowitz M, Solounias N. Embryology, comparative anatomy, and congenital malformations of the gastrointestinal tract. Edorium J Anat Embryo 2016;3:39–50. |
|
Abstract
|
|
Evolutionary biology gives context to human embryonic digestive organs, and demonstrates how structural adaptations can fit changing environmental requirements. Comparative anatomy is rarely included in the medical school curriculum. However, its concepts facilitate a deeper comprehension of anatomy and development by putting the morphology into an evolutionary perspective. Features of gastrointestinal development reflect the transition from aquatic to terrestrial environments, such as the elongation of the colon in land vertebrates, allowing for better water reabsorption. In addition, fishes exhibit ciliary transport in the esophagus, which facilitates particle transport in water, whereas land mammals develop striated and smooth esophageal musculature and utilize peristaltic muscle contractions, allowing for better voluntary control of swallowing. The development of an extensive vitelline drainage system to the liver, which ultimately creates the adult hepatic portal system allows for the evolution of complex hepatic metabolic functions seen in many vertebrates today. Human digestive development is an essential topic for medical students and physicians, and many common congenital abnormalities directly relate to gastrointestinal embryology. We believe this comprehensive review of gastrointestinal embryology and comparative anatomy will facilitate a better understanding of gut development, congenital abnormalities, and adaptations to various evolutionary ecological conditions.
| |
|
Keywords:
Anatomy education, Digestive, Embryology, Gastrointestinal tract
| |
|
Introduction
| ||||||
|
The gastrointestinal tract is one of the earliest organ systems to form in the human embryo; it develops as a diverticulum off the yolk sac. The most primitive fishes exhibit a tubular digestive tract, functioning proximally in transport and distally in nutrient breakdown and absorption. The expansion of this early gut tube into an intricate digestive system involving accessory organs and intestinal rotation reflects the evolution of more complex dietary habits. Several features of gastrointestinal development also demonstrate the transition from water to land organisms. A recent article reviewed head and neck embryology with links to evolutionary anatomy, to provide context to the morphology of primitive structures, and explain complexities in head development [1]. Similarly, we believe the study of digestive comparative anatomy puts the embryology and modern anatomy into perspective, and facilitates a deeper comprehension of the human gastrointestinal tract. Feeding to obtain nutrients, such as pre-synthesized amino acids, carbohydrates, and smaller biomolecules is an essential process common to all multicellular organisms. It is crucial to absorb organic molecules and water, and subsequently break down the materials to be used in various cellular processes. There are three types of multicellular organisms: fungi, plants, and animals. Fungi are composed of filaments that invade decomposing materials such as fruits or carcasses. They absorb the nutrients at the site and incorporate them into their own cells [2]. Plants rely on solar energy, which is converted to usable nutrients via chlorophyll molecules. They also obtain minerals and water from their roots that penetrate the surrounding soil [3] [4]. Animals ingest food, which is subsequently processed by a tubular digestive system. The food particles are broken down mechanically and chemically by digestive enzymes and intestinal bacteria. Both absorption and digestion take place in the intestines, and wasted bacteria and unused food material is excreted. Comparative anatomy and evolutionary biology provide context to the development of early digestive structures, however these courses have been lost in the pre-medical and medical curricula [5]. A key concept in anatomy involves relating form to function; comparative anatomy demonstrates how structural adaptions can meet environmental or changing physiologic needs. A current recommendation by the American Association of Medical Colleges is to incorporate evolutionary biology in medical education [6]. Many articles argue the importance of Darwinian medicine in training future physicians [7] [8][9]. Evolutionary biology teaches medicine at the molecular level, for example demonstrating the virulence of viruses via genetic mutations, and at the epidemiological level, such as relating malarial resistance to the sickle-cell trait carrier by natural selection [10] [11]. Studies in North America and in the UK showed a need for evolutionary biology to be incorporated into standard medical education courses [12][13]. In this article, we summarize the key embryological events in human gastrointestinal development, while providing connections to comparative digestive anatomy. The evolutionary history of the gut tube puts the modern anatomy and development into context, and enriches the medical student knowledge of digestive adaptations by relating structure to function. We also describe important congenital anomalies of the gastrointestinal tract; an understanding of these anomalies helps reinforce knowledge of the normal embryology. We believe this comprehensive review of gastrointestinal embryology and comparative anatomy will enable medical students to better understand gut development and adaptations to various evolutionary ecological conditions. | ||||||
|
The General Development Of The Gastrointestinal Tract
| ||||||
|
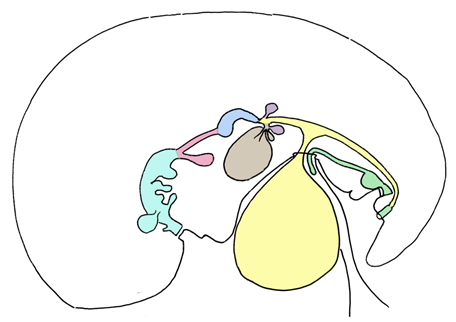
During third to fourth week of development, the embryo folds and the innermost endodermal layer forms the gut tube, which communicates ventrally with the yolk sac (Figure 1). The gut forms as a diverticulum off of the yolk sac. The gut tube is divided regionally into the foregut, which terminates cranially in the oropharyngeal membrane, midgut, and the hindgut, which terminates caudally in the cloacal membrane. The oropharyngeal and cloacal membranes are the only two regions in the body where ectoderm directly contacts endoderm. These membranes eventually rupture, making the gut tube a patent pathway proximally and distally. As folding and gut tube lengthening continue, the ventral connection between the midgut and yolk sac narrows and elongates to form the vitelline duct. The endodermal gut tube is surrounded by lateral plate splanchnic mesoderm, which forms the lamina propria, submucosal, and mucosal layers of the gastrointestinal tract. The splanchnic mesoderm also forms the smooth muscles of the intestines and of the lower esophagus. Neural crest cells migrate to the developing gut tube and form the enteric nervous system. The autonomic (parasympathetic and sympathetic) innervation is also formed, largely by neural crest, and connects the gastrointestinal system to the brain. Therefore, the gut tube is controlled by two nervous systems [14]. The gut tube is surrounded by the splanchnopleure, which ultimately forms the visceral peritoneum. A broad layer of mesenchyme initially attaches the abdominal gut tube to the body wall. This attachment subsequently thins and forms the dorsal mesentery. The gut tube becomes suspended in the peritoneal cavity, and the viscera therefore are intraperitoneal. These intraperitoneal structures are suspended in mesenteries, but are still mobile. Retroperitoneal structures are organs deep to the abdomen, or posterior to the peritoneal cavity. Primarily retroperitoneal structures originate behind the peritoneal cavity, and are never suspended; these include the kidneys, adrenals, ureters, and bladder. Some regions of the gut tube that develop initially within the peritoneal cavity are rotated and fuse to the posterior body wall; these are termed "secondarily retroperitoneal" and include the duodenum, pancreas, ascending colon, and descending colon [15]. Evolutionary link The regional delineations of the foregut, midgut, and hindgut are not common to all species. In the more primitive form, such as in sharks and certain fishes, there is minimal anatomical distinction between the small and large intestines, and often there is no discrete out pouching to form the stomach. The pylorus, however, which delineates the transition from stomach to intestines, is common to most vertebrates. Primitively, the gut tube regions proximal to the pyloric constriction functioned in food transport from the mouth, and regions distal to the pylorus were responsible for digestion and absorption. The foregut evolves to play a larger role in nutrient breakdown and food storage, and the stomach becomes pivotal for chemical and mechanical digestion [17]. | ||||||
|
| ||||||
|
| ||||||
|
Greater Sac, Lesser Sac, And Mesenteries
| ||||||
|
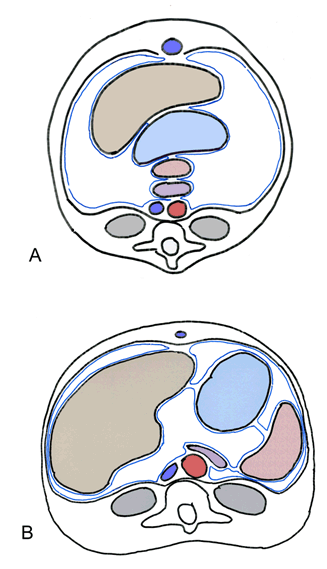
Initially, the pancreas, spleen, stomach, and liver are aligned dorsoventrally in the median plane, and are connected by ligamentous or mesenteric attachments. The pancreas and spleen are joined within the dorsal mesogastrium, the spleen and stomach are connected by the gastrolienal ligament, the stomach and liver are joined by the hepatogastric ligament, and the liver connects to the ventral wall with the falciform ligament (Figure 2). This line of organs divides the abdominal cavity into two initially equal areas. However, as abdominal organs enlarge and rotate, the dorsal mesogastrium is repositioned and creates a small space behind the stomach termed the lesser peritoneal sac. The remainder of the peritoneal cavity, comprising the majority of the abdomen becomes the greater peritoneal sac; the two spaces are connected via the epiploic foramen. The epiploic foramen represents the central point of embryonic rotation [18]. The stomach attaches to the posterior body wall via the dorsal mesogastrium. The double-layered dorsal mesogastrium extends caudally during rotation of the stomach, and fuses with the double-layered mesentery of the transverse colon to create the greater omentum. The greater omentum is thus composed of four layers of visceral peritoneum. The lesser omentum extends from the stomach and duodenum to the liver and includes the hepatogastric ligament, which joins the liver with the stomach, and the hepatoduodenal ligament, which connects the liver to the duodenum. The hepatoduodenal ligament contains the portal triad (bile duct, portal vein, and hepatic artery) [19]. Evolutionary link | ||||||
|
| ||||||
|
| ||||||
|
Esophagus
| ||||||
|
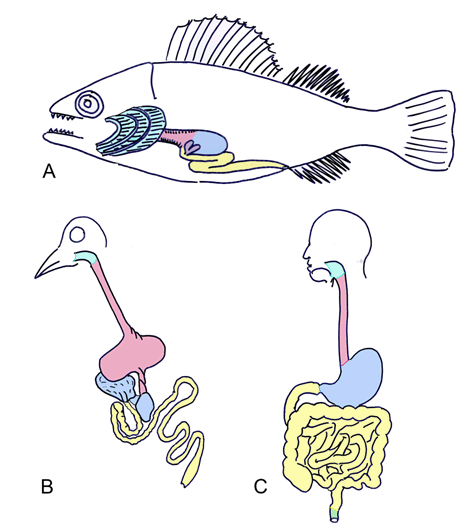
The esophagus is the cranial-most structure formed from the foregut. The respiratory diverticulum forms on the ventral surface of the developing esophagus; this will further differentiate to form the trachea, bronchi, and lungs. The respiratory diverticulum is separated from the cranial foregut by the tracheoesophageal septum. The esophagus is initially a short structure that progressively grows to span the length of the thoracic cavity. The inner wall of the esophagus is comprised of endoderm. The musculature of the upper esophagus is striated muscle, derived from branchial arch mesoderm, and the musculature of the lower esophagus is smooth muscle, derived from splanchnic mesoderm [21]. Evolutionary link | ||||||
|
| ||||||
|
| ||||||
|
Stomach And Duodenum
| ||||||
|
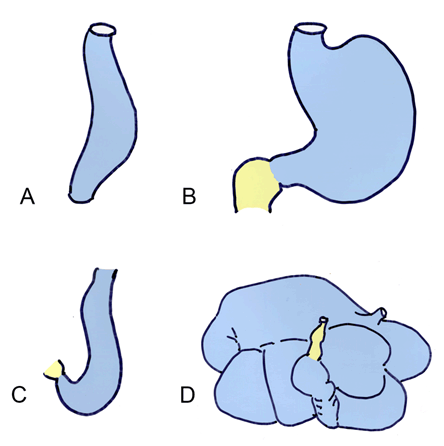
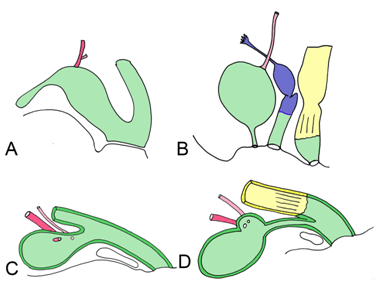
The stomach forms as a dilatation of the foregut. The developing stomach initially connects to the ventral body wall via the septum transversum. This eventually thins and forms the ventral mesentery. Around fifth week, the stomach wall grows asymmetrically; the dorsal wall grows faster and creates the greater curvature, and the ventral wall forms the lesser curvature. During weeks seven and eight, the stomach rotates 90 degrees around a longitudinal axis, bringing the greater curvature to the left and the lesser curvature to the right (Figure 4). This rotation creates the C-shaped loop characteristic of the duodenum, and moves it towards the dorsal body wall, with which it fuses to form the lesser sac. The duodenum delineates the caudal portion of the foregut and cranial portion of the midgut, and is thus supplied by both the celiac and superior mesenteric arteries [22]. Evolutionary link | ||||||
|
| ||||||
|
| ||||||
|
Liver And Gallbladder
| ||||||
|
The liver forms as a diverticulum from the foregut. Liver hepatoblasts differentiate into the hepatocytes, bile canaliculi, and hepatic ducts. The connection between the liver and duodenum narrows during development, forming the bile duct. The liver gains its hematopoietic function via migrating stem cells from the yolk sac, which is the initial site of hematopoiesis in the embryo. The liver loses its hematopoietic function shortly after the birth of the fetus, at which time the bone marrow becomes the primary site of red and white blood cell production. The liver is supplied by the vitelline vein, which brings nutrient rich blood from the yolk sac. The vitelline vein forms the adult portal vein, which drains the digestive tract. The majority of the liver is endodermal in origin, with mesodermal stromal support from the septum transversum and splanchnic mesoderm. The liver parenchyma is penetrated by branches of the vitelline and umbilical veins, forming the hepatic sinusoids. The gallbladder and cystic duct develop from the cystic diverticulum, which forms from the bile duct. The cystic duct and bile duct merge to form the common bile duct, which opens into the major duodenal papilla [21]. Evolutionary link | ||||||
|
| ||||||
|
Pancreas
| ||||||
|
Like the liver and gallbladder, the dorsal and ventral buds of the pancreas form as diverticula from the duodenum. The ventral bud forms the uncinate process and the caudal part of the head, and migrates posteriorly around the duodenum to fuse with the dorsal bud, which forms the cranial part of the head, as well as body, and tail of the pancreas (Figure 5). This migration occurs concurrently with the posterior rotation of the duodenum. The ductal systems of both buds fuse, and the ducts from the dorsal bud drain into the ventral ducts, creating the main pancreatic duct. This drains into the duodenum at the ampulla of Vater, along with the common bile duct. The pancreatic endoderm differentiates into cells that perform the many functions of the pancreas. The exocrine (acinar) cells function in digestion, the ductal cells function in transport, and the endocrine cells produce hormones such as insulin, glucagon, and somatostatin. The endocrine cells aggregate into islets, which become functional around the fifth month of development [14]. Evolutionary link | ||||||
|
| ||||||
|
| ||||||
|
Rotation Of The Midgut
| ||||||
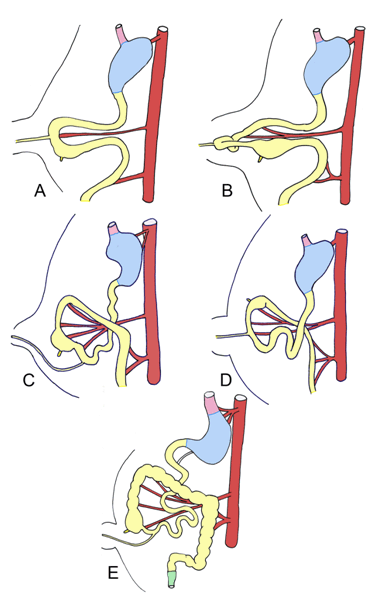
|
Around week 5, the midgut undergoes rapid elongation, and forms a hairpin loop around the superior mesenteric artery termed the primary intestinal loop (Figure 6). The cranial limb of the loop creates the jejunum, and parts of the duodenum and ileum, and the caudal limb of the loop develops into the cecum, appendix, ascending colon, and parts of the ileum and transverse colon. As the liver rapidly expands in size, the abdominal cavity becomes temporarily too small to accommodate the proliferating intestinal loop, and the intestinal loop thus herniates into the umbilicus. The midgut rotates around the superior mesenteric artery 90 degrees counterclockwise in the umbilicus, and another 180 degrees as the elongated intestines fall back into the abdomen around week 10. The jejunum and ileum also coil and loop separately from the colon. The final midgut rotates an additional 180 degrees counterclockwise, placing the cecum at the right lower quadrant. The midgut, in total, rotates 270 degrees. The ascending and descending colon contact the dorsal body wall upon their return into the abdominal cavity, and eventually fuse, becoming secondarily retroperitoneal [15]. Evolutionary link Sharks, and some primitive fishes lengthen the intestinal surface area with spiral valves within the intestinal tube. The spiral folds of mucosa extend from one end of the intestine to the other. Vertebrates expand the absorptive capacity of the intestines by extending the mucosal surface into villi and crypts, thus increasing the surface area. In many vertebrates, the intestine itself is elongated and coiled, thus necessitating its herniation into the umbilical cord to accommodate for massive lengthening during development. In mammals and birds, this elongation and rotation can generate intestinal lengths 7–8 times the height of the entire body. The absolute length of the intestines is highly variable, and often relates both to body size and dietary habits. In frog tadpoles, which are largely herbivorous, the intestines are long and extensively coiled, allowing for better plant digestion, whereas the adult frogs after metamorphosis possess shortened intestines, as they are mainly carnivorous and don't require extensive nutrient breakdown. Herbivorous species generally have more elongated intestines, allowing for the ingestion of cellulose plant material, which necessitates more extensive digestion and larger surfaces for absorption [20]. | ||||||
|
| ||||||
|
| ||||||
|
Hindgut And Cloaca
| ||||||
|
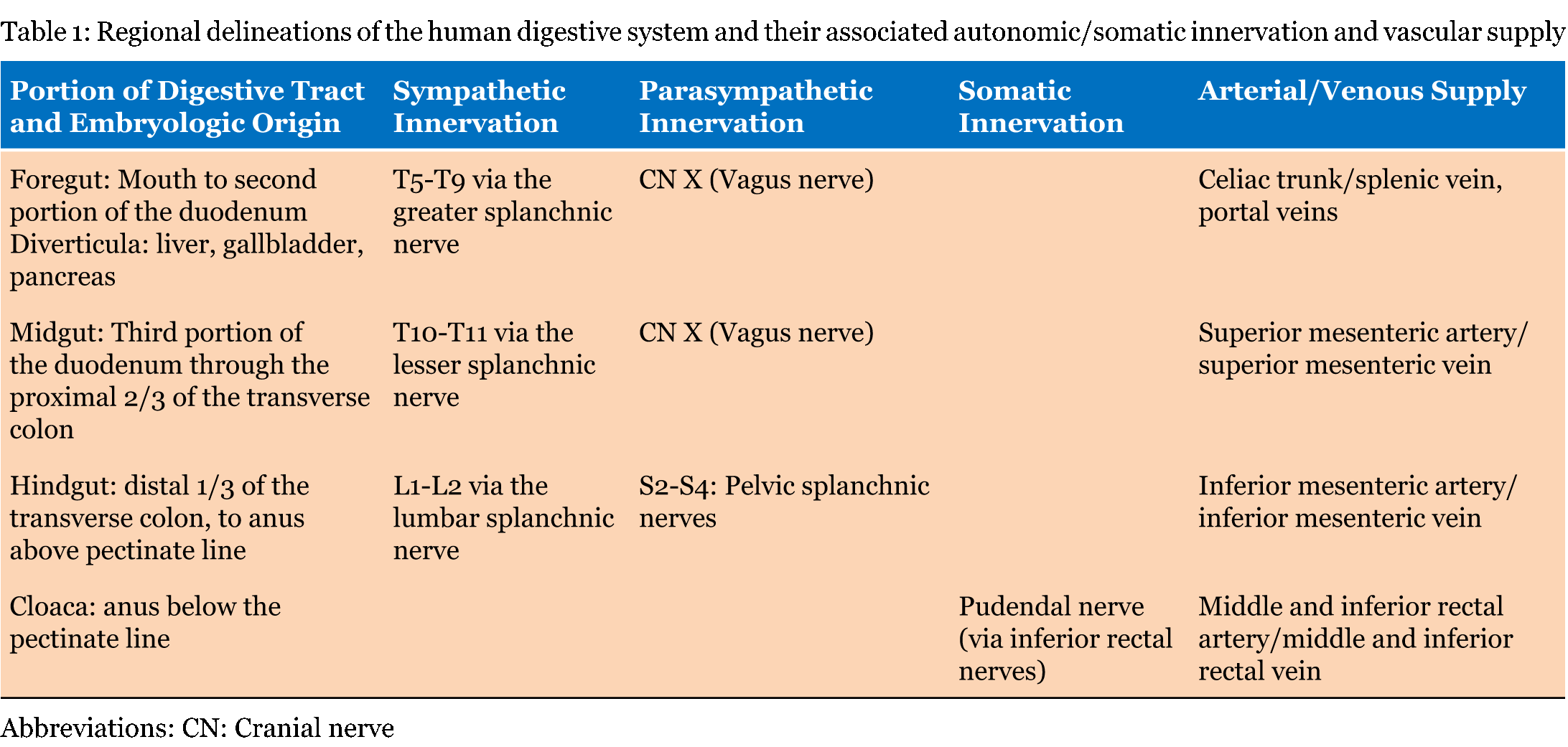
The distal-most portion of the embryonic hindgut as well as the developing urinary and reproductive tracts is continuous with the cloaca (Figure 7). The cloaca forms the anorectal canal posteriorly and urogenital sinus (future bladder, urethra, prostate, and vestibule of the vagina) anteriorly. The anus and urogenital sinus become separated by the urorectal septum, which forms the perineal body [21]. The cloacal membrane is endodermal dorsally and ectodermal ventrally, with no mesoderm in between. The membrane ruptures, allowing the anal canal to open to the exterior, thus making the gastrointestinal tract a patent pathway. In the anus, ectodermal cells of the anal membrane proliferate and temporarily plug the lumen. The membrane obliterates around week 8, and its location is marked by irregular mucosal foldings, forming the pectinate line. Anything proximal to this is formed from the endoderm of the hindgut, and distal to the pectinate line is formed by the ectoderm of the cloaca. The distal third of the anal canal is referred to as the proctodeum. As the distal and proximal anal canal have different embryologic origins, they are supplied by separate vasculature and innervation (Table 1) [14]. Evolutionary link Several groups of animals, including amphibians, reptiles, and birds, maintain the distal connection between the urinary, genital, and gastrointestinal tracts as a cloaca. The formation of a cloaca at some point during development is common to all vertebrates, however in most mammals, this distal cavity is partitioned into the anorectal canal and urogenital sinus by the urorectal septum. This septum creates separate external openings for the gastrointestinal and genitourinary tracts. In monotremes, such as the platypus, the subdivision of the cloaca is partially completed, and a fold marks the delineation between but does not fully separate the distal intestines from the distal urinary and reproductive tracts [17]. | ||||||
|
| ||||||
| ||||||
|
Vasculature
| ||||||
|
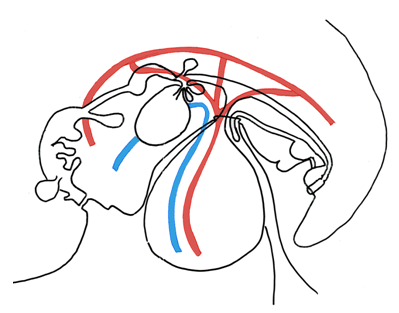
The yolk sac is initially a large and major embryonic structure, and it contains its own blood supply arising from the dorsal (abdominal) aorta. The gastrointestinal tract and associated organs derive from the yolk sac, thus the vitelline arteries persist in the adult to supply the digestive organs. The vitelline arteries arise from the aorta caudal to the diaphragm, and form in the adult the celiac artery (foregut), superior mesenteric artery (midgut) and inferior mesenteric artery (hindgut). During early development, these arteries bring deoxygenated blood from the heart towards the yolk sac [22]. The embryo has three venous systems: the vitelline system, the umbilical system, and the cardinal system. The umbilical and vitelline veins bring blood from the placenta and yolk sac, respectively, into the liver and heart. The vitelline and umbilical systems contribute to the ultimate venous drainage of the developed gastrointestinal tract. The vitelline veins traverse the developing liver and create the liver sinusoids. Around the third month, the left vitelline vein regresses, and the right vitelline vein drains the entirety of the gastrointestinal tract and diverticula. The vitelline venous system creates the portion of the inferior vena cava between the liver and heart, portal vein, splenic vein, and the veins draining the gastrointestinal tract, including the superior mesenteric vein and inferior mesenteric vein (Figure 8) [14]. The umbilical veins drain oxygenated blood from the placenta to the embryonic heart. The right umbilical vein ultimately regresses, and the left umbilical vein persists as the ductus venosus. The ductus venosus is an anastomosis between the inferior vena cava and umbilical vein, allowing oxygenated blood to bypass the liver and drain directly into the right atrium. This channel is closed after birth, as the placenta no longer supplies oxygenated blood, and creates the ligamentum venosum [21]. Evolutionary link | ||||||
|
| ||||||
|
Congenital Abnormalities
| ||||||
|
Tracheoesophageal fistula Pyloric stenosis Annular pancreas Intestinal malrotation Midgut volvulus Meckel's diverticulum Umbilical hernia Omphalocele Gastroschisis Imperforate anus Hirschsprung's disease | ||||||
|
Conclusion
| ||||||
|
Studying evolutionary anatomy helps put human embryology into perspective; the concepts help explain why humans develop the way we do, rather than just memorizing how. Comparative anatomy helps link form to function, and leads to an overall deeper understanding of the embryology and anatomy, and an appreciation for the complexity of other animals. Studying congenital anomalies helps reinforce embryological concepts, and learning these anomalies in their developmental context facilitates long-term retention of knowledge. We believe reviews of each system of embryology supplemented with the evolutionary anatomy and associated congenital anomalies will help students better understand human development. | ||||||
|
Acknowledgements
| ||||||
|
We thank the Department of Anatomy and the medical students of New York Institute of Technology College of Osteopathic Medicine. We thank our collaborators Bennett Futterman, Brooke Littlefield, Michelle Annabi, Kristen Farraj, and Samantha White. | ||||||
|
References
| ||||||
| ||||||
|
Suggested Reading
| ||||||
|
||||||
|
[HTML Abstract]
[PDF Full Text]
|
|
Author Contributions
Melinda Danowitz – Substantial contributions to conception and design, Drafting the article, Revising it critically for important intellectual content, Final approval of the version to be published Nikos Solounias – Substantial contributions to conception and design, Drafting the article, Revising it critically for important intellectual content, Final approval of the version to be published |
|
Guarantor of submission
The corresponding author is the guarantor of submission. |
|
Source of support
None |
|
Conflict of interest
Authors declare no conflict of interest. |
|
Copyright
© 2016 Melinda Danowitz et al. This article is distributed under the terms of Creative Commons Attribution License which permits unrestricted use, distribution and reproduction in any medium provided the original author(s) and original publisher are properly credited. Please see the copyright policy on the journal website for more information. |
|
|