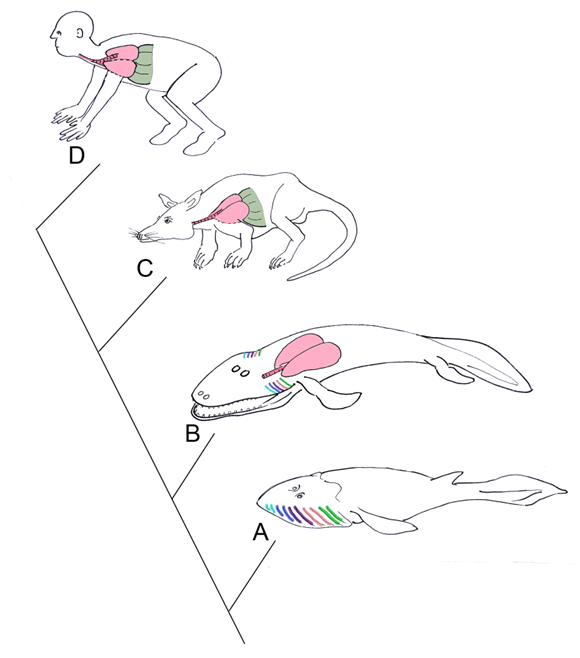| Table of Contents | |
|
Review Article
| ||||||
| Embryology and evolutionary history of the respiratory tract | ||||||
| Samantha White1, Melinda Danowitz1, Nikos Solounias2 | ||||||
|
1Medical Student, Anatomy, New York Institute of Technology College of Osteopathic Medicine, Old Westbury, NY, USA.
2Professor, Anatomy, New York Institute of Technology College of Osteopathic Medicine, Old Westbury, NY, USA. | ||||||
| ||||||
|
[HTML Abstract]
[PDF Full Text]
[Print This Article]
[Similar article in Pumed] [Similar article in Google Scholar] |
| How to cite this article |
| White S, Danowitz M, Solounias N. Embryology and evolutionary history of the respiratory tract. Edorium J Anat Embryo 2016;3:54–62. |
|
Abstract
|
|
Human respiratory embryology and anatomy often reflects the evolutionary transformation from primitive breathing apparatuses. The gills of fishes are invested with vasculature, muscles, cartilages and nerves, and function in pumping water to facilitate gas exchange. As tetrapods evolve air-breathing respiratory structures, the gills lose their breathing function. However, the associated arteries, veins, nerves, musculature, and cartilaginous support become integrated into the pharynx and head. In the Tiktaalik, a popular proposed transitional species between fishes and tetrapods, both gills and lungs are present. Variations in the anatomy of the larynx allow for differing methods of sound production between birds, reptiles, and mammals, and the changing position of the larynx in humans represents feeding mechanisms in infants, and voice production in adults. Comprehension of the normal embryologic development also facilitates a deeper understanding of congenital anomalies. The respiratory tree originates as a diverticulum off of the proximal endodermal gut tube; failed septation between the lung buds and digestive tract results in an anomalous respiratory/esophageal connection seen in tracheoesophageal fistulas. Combining key features of human lung embryology with comparative respiratory anatomy reinforces the relationship between structure and function, and will facilitate a deeper comprehension of lung development.
| |
|
Keywords:
Anatomy education, Embryology, Evolutionary biology, Gills, Lung, Respiratory
| |
|
Introduction
| ||||||
|
The study of comparative anatomy and evolutionary biology puts context to human morphology and development. Unfortunately, these courses are seldom taught in United States pre-medical and medical school curricula [1]. Comparative anatomy reinforces key concepts such as relating form to function, therefore, putting morphology into an evolutionary context. Evolutionary biology relates to medical education both at a gross and molecular level. Resistance to antibiotics is a problem plagued by hospitals worldwide; bacteria gain this virulence through natural selection [2]. The gastrointestinal tract undergoes substantial elongation in terrestrial vertebrates, allowing for enhanced water reabsorption, therefore, reflecting the evolutionary transition from aquatic to land environments [3]. Even at the epidemiologic level, evolutionary biology explains how the sickle-cell trait became so prevalent in African countries, as this gene mutation protects against malaria. A recent review of human head embryology demonstrated how evolutionary biology fosters a deeper student understanding of head and neck development [4]. Scientists and medical educators recognize the need for comparative and evolutionary biology to be incorporated into the training of future physicians [5] [6]. The evolution of a respiratory system accommodating life on land from the gills of aquatic organisms is an essential step allowing for the development of land mammals. Without a breathing apparatus that functions outside the water, terrestrial animal life would not exist. The incorporation of evolutionary biology allows for a broader and deeper understanding of respiratory tract development by emphasizing the relationship between structure and function. In this article, we review human lung embryology and provide links to the key evolutionary events in respiratory development, therefore, providing context to the morphology of the embryologic and adult structures. | ||||||
|
Pharynx
| ||||||
|
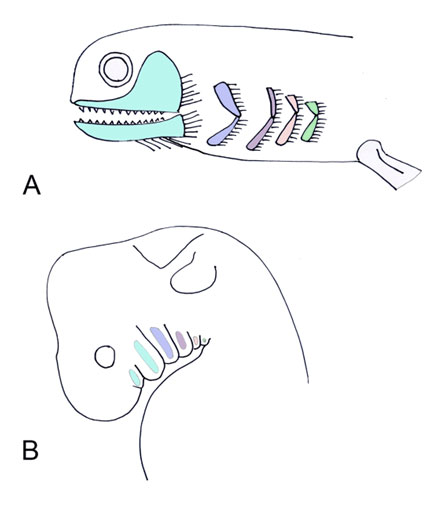
The pharynx is a complex structure that connects the oral cavity to the proximal respiratory and digestive systems. The pharynx develops from a unit system of arches, clefts, and pouches. The clefts are positioned externally, and the pouches internally, with the corresponding arch in between. Each arch derives from head mesenchyme, and includes an associated artery, branchiomeric cranial nerve, muscles, cartilages, and neural crest cells. There are five pharyngeal arches, each of which include a cranial nerve (Figure 1). The cranial nerves of the pharyngeal arches (CN V, VII, IX, and X) have mixed modalities, including motor and sensory roles. These nerves also have glandular secretory functions, as they innervate the associated endocrine structures derived from their respective pharyngeal pouch. All muscles derived from a pharyngeal arch are innervated by the respective cranial nerve; for example, the muscles of facial expression derive from the second pharyngeal arch, and are therefore innervated by CN VII. The arches and cleft-pouch units comprise the wall of the pharynx, and create the pharyngeal floor and mucosal surface of the tongue [7]. Evolutionary link: Gills As lungs evolve, the importance of the gills decrease, and the pharynx consequently decreases in size. Sarcopterygians are bony fishes that possess bones and musculature in their paired fins that resemble the limbs of tetrapods. Lungs are found in sarcopterygians , likely as an adaptation for survival in arid environments. Lungs would have enabled these fishes to leave small inhabitable ponds, to find larger bodies of water. The sarcopterygians evolved during the Devonian period (400 million years ago) during frequent seasons of drought, therefore, their survival required the presence of air breathing respiratory structures. Amphibians are transitional in respiratory evolution; the larva that develops underwater possesses external gills, whereas the adult possesses simple lung forms, allowing for air breathing on land [9]. The Tiktaalik is likely the species representing the transition from fish to amphibians, as it shares anatomical features seen in primitive fish as well as early tetrapods. Tiktaalik possesses both gills and lungs (Figure 2). Many modern fishes utilize gills for respiration, and possess a swim bladder for buoyancy in changing water depths. The majority of tetrapods, however, lose their functioning gills, and utilize the lungs for air breathing. The original gill structures become incorporated into the head and neck of land dwellers as pharyngeal arches with associated muscles, nerves, and vasculature, but loose their respiratory function. The lungs of Tiktaalik likely allowed the animal to spend periods of time out of the water, or in water with poor oxygen content. This animal also possesses a full set of ribs, assisting with air breathing, and providing support to its body. Typical fishes possess dorsal and ventral sets of ribs with a different orientation of intercostal muscles from tetrapods; fishes do not rely on ribs for breathing, but rather respire with water flowing through their gills [10]. | ||||||
|
| ||||||
|
| ||||||
|
| ||||||
|
| ||||||
|
Larynx, Trachea, and Bronchi
| ||||||
|
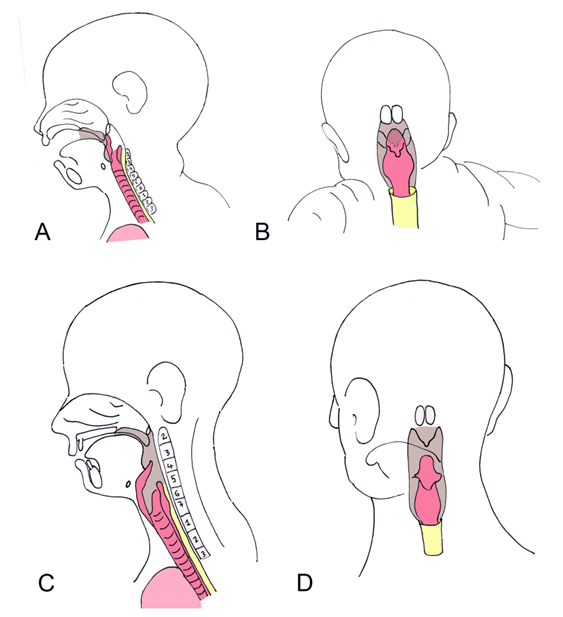
The complex respiratory tree originates as an outpouching, termed the respiratory diverticulum, or lung buds. This forms as a diverticulum at the proximal end of the endodermal foregut (future esophagus). The inner surface lining of the larynx therefore is endodermal. The 4th and 6th pharyngeal arches form the laryngeal cartilaginous support, including the thyroid, cricoid, and arytenoid cartilages. The musculature of the pharynx and larynx also derives from the 4th and 6th arches, and are, therefore, innervated by branches of the vagus nerve (CN X). The proximal stem of the respiratory diverticulum forms the trachea and larynx, whereas the distal portion forms the bronchi, bronchioles, and eventually, alveoli [11]. Evolutionary link The larynx is an enlarged cartilaginous vestibule that is found in land tetrapods. Although many tetrapods are voiceless, the larynx is often used for sound production. Frogs produce sounds using tracheal ridges. Lizards and snakes produce hissing noises by pushing air violently through the glottis. Most mammals produce voice through a pair of laryngeal vocal cords, which are ridges of elastic tissue and ligaments that are stretched across the larynx. Birds possess a larynx but lack vocal cords; their voice production takes place through the syrinx. The syrinx is a specialized structure positioned further down the trachea than the larynx, and is positioned at the division into the two major bronchi [8]. With the exception of the sarcopterygii lungfish that possess lungs extending directly from the pharynx, many lung bearing species have a midline passageway as the trachea. The trachea in tetrapods extends from the larynx to the bifurcating primary bronchi. The trachea is lined with ciliated epithelium, mucous and glandular cells, and is strengthened by the surrounding cartilaginous rings [3]. | ||||||
|
| ||||||
|
| ||||||
|
Development and Maturation of the Lungs
| ||||||
|
The lung develops as a composite of endodermal and mesodermal tissues, and matures in 5 broad stages (Table 1). The endoderm of the respiratory diverticulum gives rise to the mucosal lining of the respiratory tree and to the endothelial cells of the alveoli. The supporting musculature and cartilages that surround the bronchi, as well as the visceral pleura that covers the lungs derive from splanchnopleuric (lateral plate) mesoderm. Around the end of the 4th week, the laryngotracheal diverticulum forms two lateral outpouchings, termed the lung buds. These lung buds branch into the bronchi and bronchial tree during the pseudoglandular and canalicular stages of development [13]. The primary bronchial buds undergo approximately 16 rounds of branching, therefore forming the respiratory tree (Figure 4). The branching lung endoderm becomes invested in the surrounding mesenchyme, and cellular interactions between the endoderm and mesoderm regulate the branching. The first round of branching of the primary bronchial buds around week 5 yields three secondary bronchial buds on the right and two on the left; these secondary buds give rise to the lung lobes. This round of branching forms the three lobes of the right lung and two lobes of the left lung. The secondary bronchial buds undergo an additional 14-16 rounds of branching, and form the respiratory tree through the terminal bronchioles by the end of the canalicular stage. During the sacular stage, the bronchioles divide into the respiratory bronchioles and eventually into the terminal sacs (also termed primitive alveoli). Around week 36, the terminal sacs become invested in surrounding capillaries, allowing for future gas exchange from the respiratory to the vascular system. The alveoli continue to mature even into childhood [14]. The type II alveolar cells produce a liquid substance termed surfactant, which lines the inner surface of the alveolar endothelium. The surfactant increases the surface tension and prevents the alveolar sacs from collapsing. The developmental maturity of the lungs is a key determinant of survival in premature infants, as the lungs lack sufficient surfactant before 30 weeks, and therefore the alveoli are easily collapsible during expiration, preventing effective respiration in the newborn. During development, the fetus constantly swallows amniotic fluid produced by the kidneys. This fluid bathes the maturing lungs, and is crucial for its final maturation and expansion of alveoli [11]. There is a dual system of circulation in the lung. The lung tissue itself is supplied by the bronchial arteries, which branch off the descending aorta. The respiratory function of the lungs is supported by the pulmonary artery and branches; deoxygenated blood is transported directly to the lungs, where the blood becomes oxygenated via gas diffusion at the alveoli. The newly oxygenated blood is returned to the heart via the pulmonary vein, where it is subsequently pumped to the entirety of the body [7]. Evolutionary link: swim bladder and air sacs Birds evolve air sacs, in addition to the typical lungs, that maximize gas exchange by modifying lung ventilation. These air sacs are located in the neck, thorax, and abdomen, and are continuous with the main respiratory tract and lungs (Figure 5). They are not highly vascularized as are the lungs, and they do not directly participate in oxygen exchange during respiration. They function as bellows that create a unidirectional flow of air, which, unlike the bidirectional mammalian system, allows for more efficient oxygen diffusion into the blood. This optimization of respiration allows birds to respire at higher elevations, and also plays a key role in thermoregulation [16]. | ||||||
| ||||||
|
| ||||||
|
| ||||||
|
| ||||||
|
| ||||||
|
Diaphragm
| ||||||
|
The diaphragm originates from several embryonic structures; the septum transversum, pleuroperitoneal membranes, and body wall mesoderm. The fibrous septum transversum is originally positioned near the upper cervical somites, and therefore its innervation by the phrenic nerve (C3-C5) reflects this early positioning. The septum transversum creates the non-muscular central tendon of the diaphragm, and is the first partition between the thoracic and abdominal cavities. The bulk of the diaphragm muscle is positioned within the pleuroperitoneal membranes, and is innervated by the phrenic nerve. The peripheral rim of diaphragmatic musculature derives from body wall mesoderm, and is supplied by spinal nerves T7-T12. The crura of the diaphragm originate on the upper lumbar vertebral bodies and condense into right and left muscular bands inserting into the dorsomedial diaphragm [14]. Evolutionary link | ||||||
|
| ||||||
|
Congenital Abnormalities
| ||||||
|
Choanal atresia Laryngeal webs Laryngomalacia Vascular ring Tracheoesophageal fistula Kartagener syndrome (primary ciliary dyskinesia) Respiratory distress syndrome Congenital diaphragmatic hernia Pulmonary hypoplasia and oligohydramnios | ||||||
|
Conclusion
| ||||||
|
An understanding of the general evolutionary patterns of the respiratory tree puts lung anatomy into perspective, and reinforces the relationship of anatomical structure to its function. Comparative anatomy is a key subject for training physicians, which is often lacking in the medical curriculum, and it intimately relates to human embryologic development and adult anatomy. A deeper understanding of the embryologic processes also facilitates comprehension of congenital pathologies, as these anomalies reflect errors in development. We believe focused yet comprehensive reviews of human embryology supplemented with evolutionary biology and congenital abnormalities will broaden the medical student knowledge of development. | ||||||
|
Acknowledgements
| ||||||
|
We thank the Department of Anatomy at New York Institute of Technology College of Osteopathic Medicine. We also thank Wolfgang Gilliar and Jerry Balentine for their continued support and assistance. | ||||||
|
References
| ||||||
| ||||||
|
Suggested Reading
| ||||||
| ||||||
|
[HTML Abstract]
[PDF Full Text]
|
|
Author Contributions
Samantha White – Substantial contributions to conception and design, Acquisition of data, Analysis and interpretation of data, Drafting the article, Revising it critically for important intellectual content, Final approval of the version to be published Melinda Danowitz – Substantial contributions to conception and design, Drafting the article, Revising it critically for important intellectual content, Final approval of the version to be published Nikos Solounias – Substantial contributions to conception and design, Drafting the article, Revising it critically for important intellectual content, Final approval of the version to be published |
|
Guarantor of submission
The corresponding author is the guarantor of submission. |
|
Source of support
None |
|
Conflict of interest
Authors declare no conflict of interest. |
|
Copyright
© 2016 Samantha White et al. This article is distributed under the terms of Creative Commons Attribution License which permits unrestricted use, distribution and reproduction in any medium provided the original author(s) and original publisher are properly credited. Please see the copyright policy on the journal website for more information. |
|
|