| Table of Contents | |
|
Original Article
| ||||||
| Morphological and morphometric study on the early developmental stages of the Egyptian toad Bufo regularies Reuss | ||||||
| Alaa El-Din H. Sayed1, Ekbal T. Wassif1, Afaf I. Elballouz1,2 | ||||||
|
1Zoology department, Faculty of Science, Assiut University, 71516 Assiut, Egypt.
2Zoology department, Faculty of Science, Misurate University, Misurate, Libya. | ||||||
| ||||||
|
[HTML Abstract]
[PDF Full Text]
[Print This Article]
[Similar article in Pumed] [Similar article in Google Scholar] |
| How to cite this article |
| Sayed AH, Wassif ET, Elballouz AI. Morphological and morphometric study on the early developmental stages of the Egyptian toad Bufo regularies Reuss. Edorium J Anat Embryo 2016;3:1–14. |
|
Abstract
|
|
Aims:
This study aimed to examine the developmental changes in external morphology characters for some embryonic larval period of Bufo regularies from River Nile, Assuit, Egypt.
Methods: Our experiment started when tadpoles began to feed. The adapted embryos were divided into three large tanks, each with 200 embryos. Collecting of samples started from feeding age every three days until end of metamorphosis (the absorption of tail). External measurements of the early developmental stages including body height (BH), body width (BW), eye diameter (ED), forelimb length (FLL), head-body length (H-BL), hind limb length (HLL), internarial distance (IND), interorbital distance (IOD), nostril diameter (ND), distance between nariel and orbital (NOD), tail length (TAL), tail height (TH), total length (TL), tail muscle height (TMH) and body weight (BWE) were recorded. Results: The morphological description reported for each stage indicated many differences between the different developmental stages. Conclusion: This study may be considered as morphological key for the different developmental stages of the Egyptian toad tadpoles (Bufo regularies). | |
|
Keywords:
Bufo regularies, Egyptian toad, Morphology, Tadpoles
| |
|
Introduction
| ||||||
|
Bufonidae represent one of the common anuran families distributed in all parts of the world except for Antarctica. About 300 species are included in the Bufo genus. The common species in Egypt are Bufo regularies, Bufo viridis, Rana bedriagae and Ptychadena mascareniesis [1] . The knowledge of tadpole morphology is a valuable tool for different biological studies, i.e. taxonomic, ecological, bio-geographic and faunistic registers. In addition, it had been reported that the knowledge of tadpole's morphological aspects can serve as a support to understand species preservation, and information that helps understand basic and derivative aspects of these species [2]. Anuran metamorphosis consists of an early period of larval growth (premetamorphosis), a subsequent interval of continued growth and development of some adult organs (prometamorphosis) and a final period when animals cease feeding, larval organs are resorbed, and final transformation to the juvenile frog occurs (climax) [3][4]. Two approaches have described the anurans ontogeny, the first one has named life cycle events as stages; embryonic, larval, metamorphic climax and juvenile/adult [5][6]. The second approach has addressed ontogeny as metamorphosis that can be subdivided into three periods; premetamorphosis, prometamorphosis, and metamorphic climax [7] [8][9][10]. Some studies investigated that the beginning of the larval period as when feeding begins [10] while others stated that the completion of hatching is the beginning [5][6]. When comparing species, the morphological development at hatching differs between species [7][8] [10] observed that the emergence of forelimb can be used as a mark of the beginning of metamorphic climax, where in some specialized anuran larvae, the initiation of mouthpart regression and forelimb emergence may be decoupled [6] [11]. It has been reported that the anuran metamorphosis is controlled by the hypothalamo-hypophyseal-thyroid axis involving actions of several hormones [12][13][14] such as thyrotrophic releasing hormone (TRH), thyroid hormones (TH), and prolactin [4] [15] [16]. Considerable flexibility has been observed in the rates of growth and development by several authors [17][18]. Morphological characteristics are important marks to distinguishing between tadpoles of different species [19] . The external larval stages can also be identified on the basis of age, size and external morphological characters [4]. Only two works have been published on the external morphology of the early development of the tadpole of Bufo regularies by several authors [20][21] who had suggested a generalized table of 66 stages covering the period from fertilized eggs to the completion of metamorphosis. In the present works the tadpoles of the Egyptian toad Bufo regularies will be re-described, The normal table that suggested by Sedra and Michael [20] are still the source of information about these species. The aim of the present study is to identify the different developmental stages of the Egyptian toad Bufo regularies; a step toward building a morphological and morphometric key for larval stages of the Egyptian toad (Bufo regularies). This search represented the first one of our series future papers about the development of Bufo regularies tadpoles. | ||||||
|
Materials and Methods
| ||||||
|
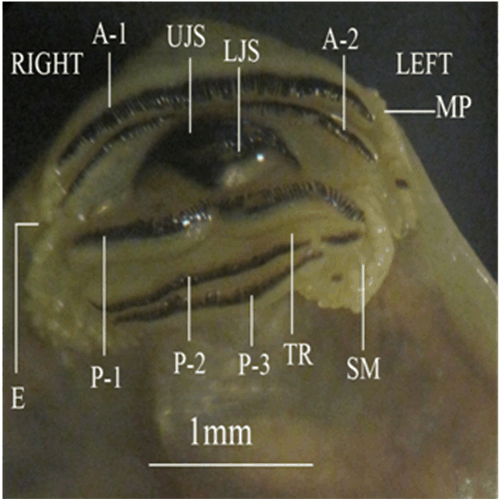
Specimen collection Experimental Setup Morphology and Morphometric measurements The external morphology of the different developmental stages were recorded including: Body Height (BH), Body Width (BW), Eye diameter (ED), Forelimb Length (FLL), Head- Body Length (H-BL), Hind Limb Length (HLL), Internarial Distance (IND), Interorbital Distance (IOD), Nostrial Diameter (ND), Distance between Narial and orbital (NOD), Tail Length (TAL), Tail Height (TH), Total length (TL), Tail Muscle Height (TMH) and Body Weight (BWE). The body parts were based on the proposals of [24] . The external morphometric measurements were described in (Figure 1a-b). Statistical analysis Ethical statement | ||||||
|
| ||||||
|
Results | ||||||
|
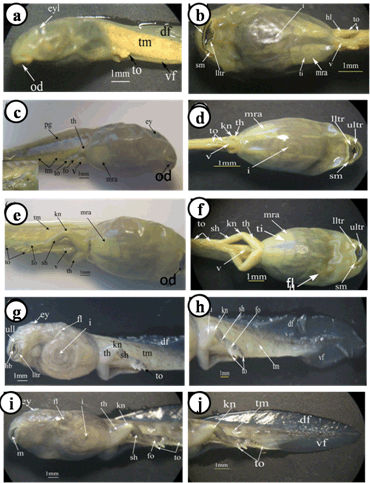
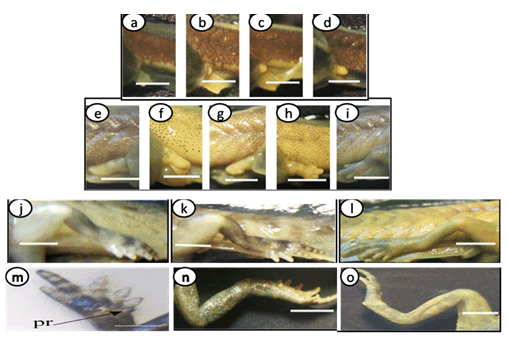
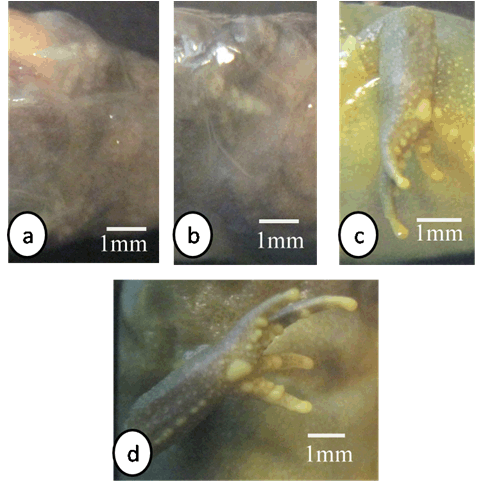
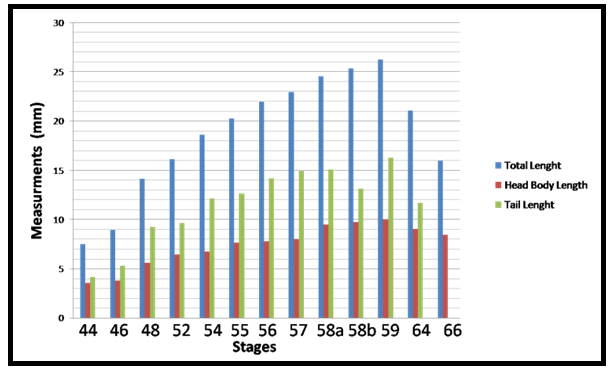
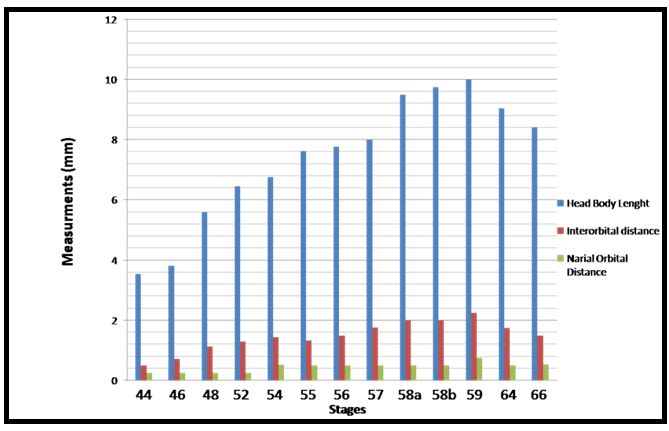
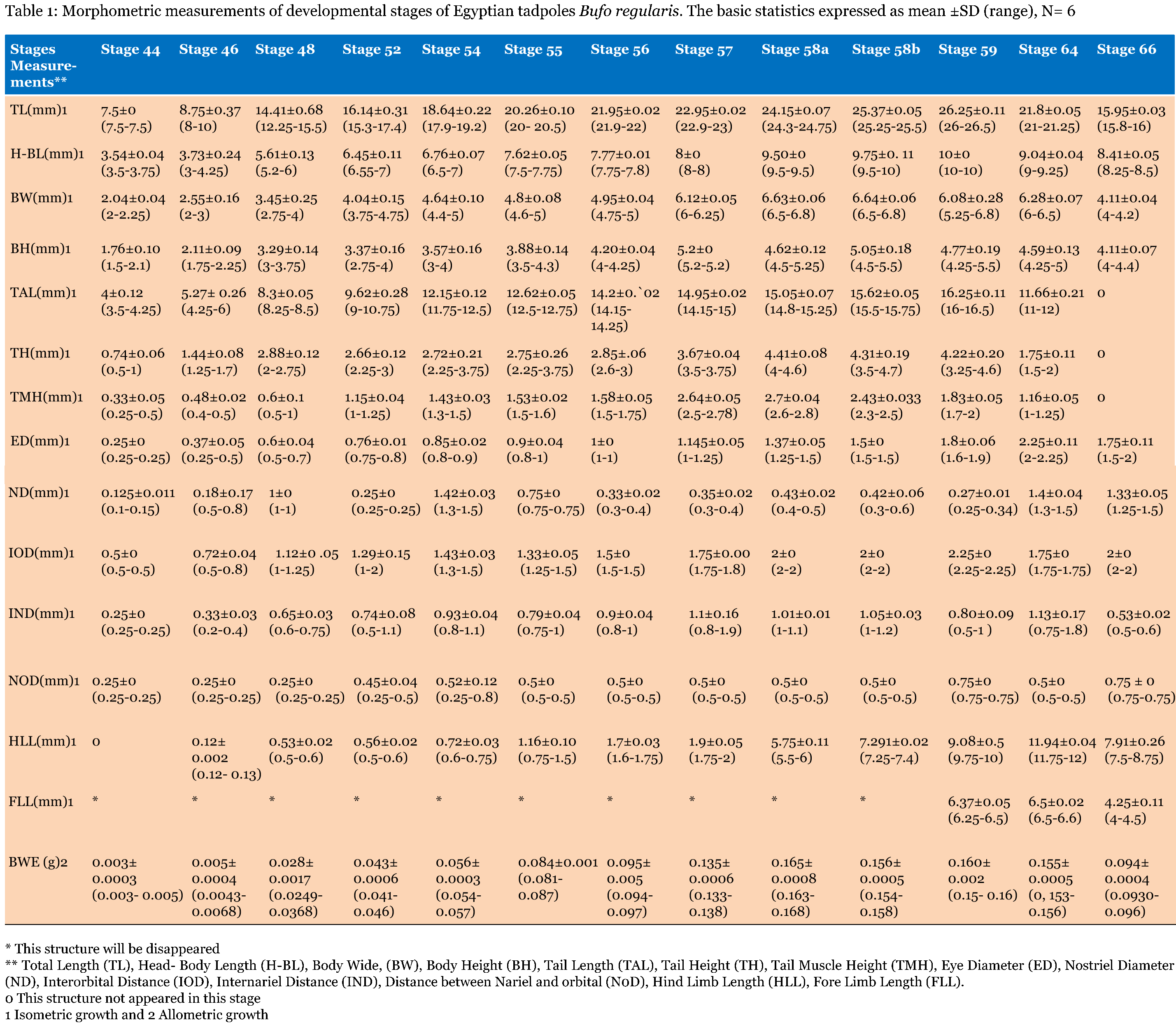
The important larval stages were differentiated on the basis of age, size and external morphological characters. Various stages of development and metamorphosis of Bufo regularies were divided into 13 stages. These were collected every three days from beginning of feeding to the end of the metamorphosis. This description was based on the normal table of [20] . The investigators described only the new structures appeared in each stage. Morphology Stage 44: Start feeding stage (TL; 7.5–7.5 mm, BWE; 0.003–0.005 g). At the beginning of feeding the body (head and abdomen) is oval in shape and pigmented dorsally, but ventrally there is no pigments. The dorsally located eyes are moderately large in size. The nares are oval in shape and positioned between the eyes and the oral disc. The protocercle tail is tapering and laterally compressed (Figure 2a-b). The pigmented tail musculature is poorly developed and gradually tapering from base to the sharp end. Unpigmented tail fin consisted of dorsal and ventral ones. They are slitghtly equal in size. The dorsal tail fin slightly smaller than the ventral one. Dorsal fin originates at body - tail junction and expanded slightly above the body. The lateral tubular vent is moderately large and directed posteriorly, entirely attached to ventral fin (Figure 2b). Coiling intestine appeared as triple U-shape ventrally. The mouth is located on the anteroventral side of the head. The small oral disc is transversally elliptical and consists of upper and lower lip. These lips carry teeth (Figure 2a&b). Stage 46: Appearance of the hind limb buds (TL; 8–10 mm, BWE; 0.0043–0.0068 g) In this stage, hind limb bud was visible for the first time as a faintly circumscribed projection in the groove between the base of the tail and body wall, (Figure 2c and 6a). Horny labial tooth rows of upper and lower oral disc were well developed. The first indication of two narrow longitudinal strands of muscle fibers of the M. rectus abdominis were observed through the transparent skin of the abdomen. Intestine is more coiling. Melanophores began progressively spread ventrally. Segmentally arranged myotomes are noticed for the tail muscles (Figure 2c-d). Stage 48: Length of hind limb bud was equal to one and half times of its diameter (TL 12.5–13 mm; BWE; 0.024–0.036 g) Length of hind limb bud was equal to one and half times of its diameter (Figure 2e and 6c). Horny teeth and sub marginal papillae very highly developed. Through the transparent skin the elongation of the two narrow longitudinal strands of muscle fibers of the M. rectus abdominis was observed (Figure 2f) . The Spiral coiling of the intestine increased, central coil of intestine still lies on the same level as that of the surrounds. Eye lens was very distinct. Spreading of blood capillaries in the dorsal fin was seen (Figure 2e-f). Ventral view of the tadpole showing a typical oral disk apparatus at this stage which consists of upper and lower labia, enclosing upper and lower serrated jaw sheath (horny beak). The lower serrated jaw sheath of the horny beak appeared as V shape and the upper one was crescent in shape. The upper and lower labial ridges had horny teeth. There are two rows of horny teeth in the upper lip; the anterior one is complete, while the posterior one is discontinuous medially. The lower lip has three continuous rows of horny teeth. The number and configuration of tooth rows was indicated by a labial tooth row formula (LTRF) that is 1: (1+1)/3 according to Sedra and Michael (1961). The labia were framed by lateral rows of marginal and sub marginal papillae (Figure 3). Stage 52: Foot paddle stage (TL; 15.3–17.4 mm; BWE; 0.041–0.046 g) Landmarks between parts of the hind limb were slightly distinguished. Distal end of hind limb buds were moderately flattened to form the foot paddle and slightly bent ventrally (Figures 2g and 6e). No interdigital indentation was noticed at the paddle margin. At the end of this stage first interdigital indentation were noticed. The fifth and fourth toe prominences were separated by a slight indentation at the margin of the foot paddle. Through the thin transparent skin well developed M. rectus abdominis and tail muscles were seen. Extremely distinctive segmentally arranged myotomes were noticed of the tail muscle. Each myotome is completely enclosed in a sheet of fibrous connective tissue (myocomma). The myotomes are >-shaped, each having an upper and lower limbs; their enclosing myocomma was also of the same shape and well visible externally (Figure 2g-h). Stage 54: Second indentation (TL; 17.9–19.2 mm, BWE; 0.054–0.05 g) The hind limbs were well-differentiated into thigh, shaft and foot. Second indentation occurred on the ventral side behind the prominence of 4th toe, which marks the 3rd toe (Figure 2i and 6g). Muscle fibers of M. rectus abdominis were more distinct forming an elongated sheet on either side of the middle line; this muscle is broad posteriorly and thinner anteriorly. Tendinous intersections were clearly observed (Figure 2j). Highly developed sub marginal papillae of the oral disc were noticed. Central coil of intestine pushed its way inward, (Figure 2i-j). Stage 55: Third indentation (TL 17–18.3 mm, BWE; 0.081–0.087 g) The margin of foot paddle became indented on ventral side, behind the prominence of 3rd toe which marks the prominence of 2nd toe. Pigments of melanophores were noticed on the thigh and shank of the hind limbs (Figures 4a and 6h). Tail musculature was strong, gradually tapering, almost reaching tail tip. Tail fins were moderately high, not extending above the body (Figure 4a-b). Stage 56: Fourth indentation (TL; 21.5–22 mm, BWE; 0.094–0.097 g) Hind limbs were highly differentiated into thigh, shank and foot, pigmentation is spreading on the upper surface of the shank as well as on the toe prominences (Figure 4c and 6i). The margin of foot paddle was indented behind the second toe demarcating the prominence of the first toe. All five toes were visible and separated from each other (Figure 4c). Through the transparent skin well developed forelimbs could be seen and were still concealed by the skin of the branchial chambers (Figure 4c & d). Stage 57: Toe web ws more distinct (TL; 22.5–23, BWE; 0.133–0.138 g) Landmarks between thigh, shank and foot were more differentiated and well distinct (Figure 4e); the margin of 5th toe web was directed toward the tip of 1st toe. Pigmentation appeared in the 4th and 5th toes along the foot. Toes were longer and all toes were separated from each others. Prehallux appeared as a small protuberance close to the base of the first toe. The fourth toe was the longest. Web clearly developed between 2–3 and 3–4 toes. Instep carrying little tubercles some of which are enlarged at bases of the second, third, and fourth toes (Figure 7). Muscle of M. rectus abdominis are divided into five pairs of transverse compartments by four pairs of tendinous intersections, the posterior three pairs of these compartments are highly developed as compared with the anterior ones, vent tube are highly distinct (Figure 4e-f). Stage 58a: Appearance of toe pads (TL; 25–25.3 mm, BWE; 0.154–0.158 g) The fourth toe was the largest and twice as long as the third toe. All toes were better differentiated and toe webs more distinct, the third and fifth finger were more or less equal to each other, fingers were stretched out in forelimb atrium which was still concealed by the skin of the operculum, horny teeth and jaws are still present. Muscle of M. rectus abdomens was still clearly indicated. Tendinous intersections were still obvious through skin of abdomen and replaced with muscle mass (Figure 4g-h) and (Figure 6j)(. Stage 58b: Cloacal tail piece reduced (TL; 24 – 24.75 mm, BWE; 0.163–0.168 g) Forelimbs were in process of eruption and visible through the skin (Figure 4i), elbows were pressing the overlying skin, palm well-built and is provided with a number of tubercles, vent tube disappear completely, cloacal tail piece reduced and tail darkening starts. The disappearance of teeth started (Figure 4i & j) and (Figure 6k). Stage 59: Both forelimbs emerge (TL; 27–27.3 mm, BWE; 0.15–0.16 g) The left forelimb had passed out throughout the wide, spout-like opening of branchial chamber (Figure 5a). About 4–5 tubercles on the ventral surface of each finger are present, the fourth toe is the longest. One of the tubercles on palm is more prominent. Instep carrying little tubercles some of which are enlarged at bases of the second, third and fourth toes (Figure 5b), Right forelimb has passed out, thus no continuity between skin of body and limbs. Head is semicircular in outline and clearly broader than trunk, all horny teeth and lower horny jaw have fallen down. External nares were nearer to the tip of the snout than eye. External nares were opened at this stage. Warts were appeared (Figure 5a). Muscle of M. rectus abdomens were faintly indicated through the relatively thick and opaque skin of abdomen (Figure 5a&b )and (Figure 6i). Stage 64: Angle of mouth reached posterior border of the eye (TL; 20–23 mm, BWE; 0.153–0.156 g) Tail decreased in size, dorsal and ventral lobes of caudal fin were highly reduced and represented by shallow apexes (Figure 5c). Mouth is wider and its corners lie at the level of the posterior border of eye lens, the angle of the mouth has reached posterior of the eye (Figure 8d). Rapid changes in the mouth and tail had been occurred. Linea Alba in the ventral side of the tadpole was noticed (Figure 5c&d) and (Figure 6n). Stage 66: Metamorphosed into toadlet (TL; 15.8–16 mm, BWE; 0.093–0.096 g) Newly metamorphosed toadlet: Tail has disappeared and is only represented by a minute knob which is no longer visible in ventral aspect (Figure 5e). The corner of the wide mouth lies slightly behind the posterior border of the eye. Parotoid glands lie nearly behind the eyes. The animal had small size. Horny jaws were fallen. Linea alba in the ventral side of the toadlet was clearly detected. M. rectus abdominis disappeared under the thick skin of the toadlet. Well-developed two pairs of legs; forelimb had four fingers and hind limb was longer than the forelimb and had five toes (Figure 5e&f and 6o). The development of forelimbs in different developmental stages of Bufo regularies was given in (Figure 7). | ||||||
|
| ||||||
|
| ||||||
|
| ||||||
|
| ||||||
|
| ||||||
|
| ||||||
| ||||||
| ||||||
|
| ||||||
|
| ||||||
|
Discussion
| ||||||
|
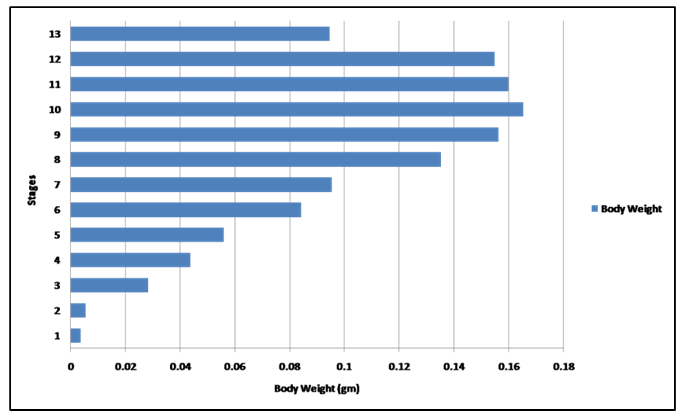
In the present study, developmental changes in external morphology characters and morphometric measurements were examined for the larval period of Bufo regularies. A developmental table consisting of 13 developmental stages from starting of feeding to complication of tail absorption every third day was recorded. To be useful, a staging table must be based on character states whose onset of development can be detected with minimal ambiguity, whose presence among individuals is stable, and, preferably, whose characteristics are quantifiable [6]. Gosner [10] suggested a generalized table of 46 stages covering the period from fertilization of the egg to completion of metamorphosis and this table has been used for the comparison of developmental stages of anurans world over. Sedra and Michael [20] had established a set of closely spaced developmental stages for the Egyptian toad Bufo regularies. These include stages numbered 1–66 which begin with the zygote stage and end with the newly metamorphosed individuals. We had used Sedra and Michael [20] staging table for identifying the stages of development of the studied tadpoles for its simplicity and general applicability. Morphology In the present study, we recorded that feeding of B. regularis tadpoles, began at stage 44. While in previous studies by Sedra and Michael [20] on the same spices noticed that at the beginning of this stage the tadpoles began route for start feeding. Gosner [10] considered the stage 25 start feeding in Bufo valliceps. In Scaphiopus bombifrons the feeding began at stage 24 [26] while in Scaphiopus intermontanus feeding began in either stage 24 or 25A [6], but in Microhyla ornate tadpole feeding began at stage 28 [27]. In Rana leptoglossa feeding began at stage 25 [4]. Hall and Larsen [6] declared that the larvae were able to begin feeding once their jaws had begun to keratinize. Sedra and Michael [20] recorded that the staging criteria associated with the development of the hind limbs which first noticeable in B. regularis during stage 46. These results were similar to the development of the hind limb bud in the Egyptian toad (B. regularis) tadpole in the present work. According to Gosner's table [10] in Bufo valliceps [10] [28] and in Rana pipiens [28] marked the initial presence of the hind limb bud as subsequent to the completion of opercullar development in stage 26. In contrast, in Rana sylvatica [29] , the bud is identifiable at the time when operculum begins development (stage 23), however, [6] noticed the first appearance of the hind limb bud in S. intermontanus during stage 24 (after opercular development had already progressed significantly but prior to completion). Similarly, Trowbridge [26] noted the first development of the hind limb bud during stage 23 in Scaphiopus bombifrons [6] described the first appearance of the hind limb bud in Scaphiopus intermontanus tadpole as an opaque swelling of tissue posterior to the abdomen and dorsal to the cloacal tailpiece. This description was in accordance with the present work. In our study in B. regularis foot paddle formation began at stage 52, when hind limb length reached one and a half times its diameter [10] had defined the foot paddle from the point when limb bud length reaches two times its diameter but formation of the foot paddle had not yet begun, and he recorded that the limb bud show as essentially bullet shaped prior to foot paddle formation (probably based on the description of stage 30 for Bufo valliceps by Limbaugh and Volpe [28], for Rana leptoglossa by Saha and Gupta [4], and for Microhyla ornata and Uperodon globulosus by Narzary and Bordoloi [18] confirmed the results of Gosner [10]. Also, they reported that stage 31 was the foot paddle shaped stage. [6] had been observed in S. intermontanus foot paddle formation began at about the point hind limb bud length was only slightly greater than its diameter. Our result was in agreement of [20] where they stated that, foot paddle formation in B. regularis occurs prior to the limb bud achieving a length of two times its diameter, but after it is one and a half times its diameter. In the present investigation, toe development in the hind limb of B. regularis tadpoles takes place from stage 52–56, where these stages takes degrees of toe development. Gosner and Taylor stated that a combination of degree of toe separation was recorded from stage 36–40 [10] [30] but, in Rana leptoglossa it had been observed that the toe differentiation occur during stages 31 to 39 [4]. In the present work, we observed first indentation between fifth and fourth toes in late stage 52. Whereas, the first indentation was noticed by Sedra and Michael [20] in stage 53 in the same species. The 2nd indentation was seen behind the fourth indentation which marks the third toe in stage 54, third indentation marks the 2nd toe at stage 55. The fourth indentation marks the 1st toe in stage 56. All five toes were visible and separated from each other. These results were similar to those described by Sedra and Micheal [20] in B. regularis. The same events were recorded by several authors [4] [18][27] in Microhyla ornata, Rana leptoglossa and Uperodon globulosus respectively, where they observed the margin of the foot paddle became slightly intended which marked the prominences of the fifth and fourth toes at stage 32, second indentation marks the 3rd toe in stage 33, third indentation marks the 2nd toe at stage 34 and the fourth indentation marks the 1st toe at stage 35. Landmarks between thigh, shank and foot with separation of all toes began at stage 56 and well differentiated at stage 57. At this stage well developed prehallux appeared as a small protuberance close to the base of the first toe. These results were similar to that described by Sedra and Micheal [20] at stages 56 and 57 in B. regularis. In our results in stage 57, we observed with the separation of all toes the fourth toe is the longest. Web clearly developed between 2–3 and 3–4 toes. Instep carrying little tubercles some of which are enlarged at bases of the second, third, and fourth toes. These results are in agreement with those of Sedra and Michael [20]. The same results were recorded in stage 39–40 by several authors [4] [18][27] in their studies. Our staging for B. regularis recorded that both forelimbs emergence and external nares are opened in late stage 59, and these results were similar to [18] [27] where they had been observed that the forelimbs emerged at (stage 42) in Microhyla ornata, also, these results were similar to Gosner and McDiarmid et al. [10] [31] where they observed that external nares open very late, after forelimbs emerge at Gosner's stage 41, and nares opened forming a rim by the nasal opening in Gosner's stage 42 [32]. In our results in stage 64, we recorded that the tail was decreased in size and the dorsal and ventral lobes of caudal fin are highly reduced and represented by shallow crests and these results were similar to stage 64 [20], also, similar to stage 44 of Microhyla ornata [27]. In Rana leptoglossa, [4] observed the tail resorption started at stage 43 with shrinking in dorsal and ventral fins. Some authors had been used the degree of tail resorption and emergence of the forelimbs as an indexes of progress in metamorphosis and they had been recorded that the tail shortening begins at about the time of forelimb emergence [7] [28]. Complete of the tail regression signals the end of metamorphosis in all tadpoles [6]. We observed the angle of the mouth reaching the posterior margin of the eye in stage 64 in B. regularis; this result was in agreement with Sedra and Michael [20]. This observation was recorded in stage 45 in Bufo valliceps and Uperodon globulosus by Gosner [10] [18] but in S. intermontanus it had been observed only in sexually mature individuals [6] . Studies on the mechanism of apoptotic cell death in the tail suggested that activation of collagenase might play an important role in the disappearance of the tail [15]. However, degradation of extracellular matrix proteins (ECM) induces proliferation, migration and differentiation of constituent cells during embryonic development [33] [34]. It has been reported that catalase inhibited programmed cell death of blastocytes and H2O2 stimulated apoptosis [35][36]. Thus, in addition to collagenase, reactive oxygen species might play critical roles in the apoptosis of cells in the tail. Our results in late stage 66, we observed completion of tail resorption and these results were similar to stage 66 [20] where they had been observed that the tail had nearly disappeared and is only represented by a minute knob which is no longer visible in a ventral aspect, and these results were similar to (stage 46) of Microhyla ornata and Uperodon globulosus [18] [27], similarly, the tail resorption was complete in stage 45 but still remained a triangular stub in Rana leptoglossa [4]. The stages of morphogenesis of the muscle rectus abdominis of the tadpoles Bufo regularies had been described from the stage of its appearance (stage 48) to the stage of its hidden under the thick skin of the tadpole at (stage 59) by Sedra and Michael [20]. In the present investigation this muscle of rectus abdominis appeared at stage 46 and obscured under the thick and opaque skin also at stage 59 (Figure 2) (Figure 4) and (Figure 5). The full developed stages of this muscle in the present investigation had been recorded from stage 56–58 but in Sedra and Michael [20] from stage 51–58. In the present work these muscles and the linea alba are well distinguished through the thin, pale toadlet skin (Figure 5f). Morphometric measurements It had been reported that the morphometric characters of tadpoles were widely used in interspecific comparison and in taxonomic studies to understand these characters and their morphology in the adult phase [37]. Also, the different morphometric variables show specific increasing or decreasing patterns in different stages [38]. Our results showed fluctuation in the TL, H-BL, BH, BW, TAL, TH and TMH, during stages 44–66, and this similar to Dey et al. and Grosjean et al. [38][39]. Previous studies have demonstrated that in the presence of predators, tail heights of tadpole become taller and overall body length become shorter [19] [40][41]. Also found that tadpoles with this increased tail height survived predation 25% more than those without increased tail heights. Similarly, Vences et al. [42] suggested that taller tail heights may be more beneficial in larger bodies of water where longer daily distances are moved than in smaller bodies of water. This is possibly due to thermoregulatory functions and possibly feeding activity. Thus the greater tail heights of Rana clamitans may be a result of their ability to exploit more habitat types as opposed to R. okalosa that only selects small bogs and seeps in Northwestern Florida [43][44][45]. Eye diameter and the inter-orbital distance was increased with age from (stage 44 to stage 64), then decreased in the late stage (stage 66), and this similar to Rana alticola [39] [40] [41][42] [43] [44] [45] [46] and these may be due to the change of the eye position from dorsal to dorsolateral position. Kruger et al. [47] suggested that dorsal eyes may face dorsally (upward), dorsolaterally, or laterally, while lateral eyes always face laterally [48]. No part of the cornea of the eye is in the dorsal silhouette when the eyes are dorsally situated. Dorsal eyes occur in benthic tadpoles in both lentic and lotic systems, while lateral eyes are most common in lentic tadpoles that spend considerable time in the water column. Also, we recorded fluctuation in the nostril diameter along the developmental stages, and this may be due to the development of respiratory system as was gills then become lungs. The interorbital distance and internarial distance also showed fluctuation along different developmental stages and this was in accordance with results of [38]. Our results showed significantly increased in hind limb length with age from (stage 46–64), then decreased in the late stage (stage 66), also, forelimb length slightly increased in stage (59 and 64) then decreased in stage 66. These results were in contrast with [38], where they recorded that the length of fore limbs continuously increased with age until tadpoles reached adult age in Microhyla ornate. Table 1, and Figure 10 showed increased in the body weight from stages 44 to 58b, then decreased in stage 59, 64, and 66, and this was similar to [49]. In contrast, [2] observed fluctuation in the body mass through development of Rhinella icterica. Decreasing in body weight was noticed from stage 59, 64 and 66 owing to the tadpoles stopped eating during the metamorphic process [20] and due to regression of the tail of the tadpoles Bufo regularies during this period and also the gradually fallen of the oral disc at this period. | ||||||
| ||||||
|
| ||||||
| ||||||
|
| ||||||
|
Conclusion
| ||||||
|
Using morphology and morphometry indicated many differences between the different developmental stages and this study may be considered as morphological key for the different developmental stages of the Egyptian toad tadpoles (Bufo regularies). | ||||||
|
Acknowledgements
| ||||||
|
This research funded by Assiut university and the Ministry Higher Education from Libyan Government. | ||||||
|
References
| ||||||
| ||||||
|
[HTML Abstract]
[PDF Full Text]
|
|
Author Contributions:
Alaa El-Din H. Sayed – Substantial contributions to conception and design, Acquisition of data, Analysis and interpretation of data, Drafting the article, Revising it critically for important intellectual content, Final approval of the version to be published Ekbal T. Wassif – Analysis and interpretation of data, Revising it critically for important intellectual content, Final approval of the version to be published Afaf I. Elballouz – Analysis and interpretation of data, Revising it critically for important intellectual content, Final approval of the version to be published |
|
Guarantor of submission
The corresponding author is the guarantor of submission. |
|
Source of support
None |
|
Conflict of interest
Authors declare no conflict of interest. |
|
Copyright
© 2016 Alaa El-Din H. Sayed et al. This article is distributed under the terms of Creative Commons Attribution License which permits unrestricted use, distribution and reproduction in any medium provided the original author(s) and original publisher are properly credited. Please see the copyright policy on the journal website for more information. |
|
|